 For ages the salamanders have been considered a class of the elementals – vehicles of the transmuting fire element which they guard and embody. Dwelling in their subtle ether, these transubstantial nature spirits were often invoked and honored via the burning of incense. Later being associated with the element nitrogen as understood within modern chemistry, the salamanders are said to be operant in the emotions of man through the liver and bloodstream (the liver is believed to be the seat of the emotional principle and assists in processing emotions in addition to its biochemical transformation roles) [1]. Salamanders (as elementals) have been assigned the cardinal direction of south, and it is maintained that they are the most powerful of the nature spirits.
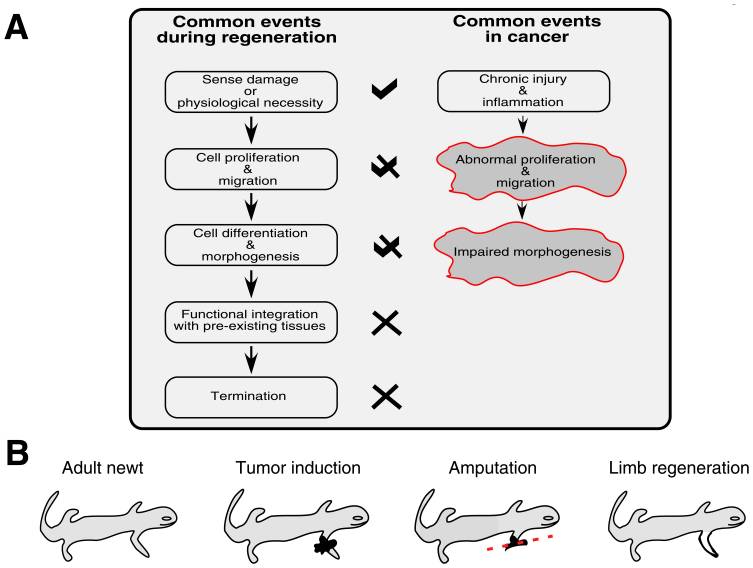
Salamanders are quite unique among vertebrates in their capacity to resist the development of cancer and regenerate limbs, and for many decades the link between regenerative ability and cancer protection has been discussed [2]. Normal tissue regeneration involves the directing of progenitor cells to an area of need, the careful proliferation and differentiation of those cells, and timely termination signals [3]. Aberrant or unregulated restoration efforts would obviously create unwanted consequences, but appropriate regeneration can actually halt and reverse the autonomous growth of malignant cells [4] [5] [6]. Because of the corollaries that exist between the events of tissue regeneration and the progression of cancerous growths, local tumors can be described as wounds that do not heal [7]. Chronic tissue stress or injury can drive ongoing inflammation which leads to an incomplete repair and restore operation that involves the misappropriating of mechanisms governing tissue mending and stem cell self-renewal – and this subversion fosters tumor maturation [8]. Because there are intracellular and extracellular factors that can regulate the self-renewal of stem cells, remedying the cancer pathology must include a reestablishment of normalcy and health to the microenvironment or terrain of the malignancy [9]. The remainder of this article will revolve around this concept as we look deeper into cancer’s idiosyncrasies. The tumor-suppressing protein p53 is frequently inactivated by mutations or viral oncogenes in human cancers, and Villiard et al. have shown that p53 signaling is necessary for limb regeneration in salamanders [10] [11]. And a related study provided evidence that more robust p53 proteins and a greater number of them confers heightened tumor resistance [12]. Ordinarily, p53 activation triggers DNA repair, apoptosis (mainly through the mitochondrial death pathway), and senescence signaling which helps to eliminate faulty cells and thwart cancerous growths [13]. Additionally, the p53 protein normally checks the self-renewal of adult stem cells, so when this protein is dysfunctional or inactive, stem cells will have less of a controlling leash, and therefore may abnormally proliferate more easily [14]. Again, the events of regeneration and tumorigenesis can be seen as two different but related paths. When the p53 protein is functioning as it should, it blocks the initial steps of regeneration from shifting toward tumorigenesis [15]. Thus, correct p53 activity helps prevent the transforming of healthy tissue restoration into unhealthy tissue malignancy. Moving forward, well-orchestrated sequences are necessary for the molding of replacement tissue in adult vertebrates to prohibit errant growth of mature cells that have dedifferentiated to fuel healing efforts. Salamanders skillfully employ dedifferentiation and transdifferentiation of mature cells to rebuild tissues that have been damaged or removed [16]. As Oviedo and Beane have explained, “in mammals chronic epithelial injury often precedes malignant transformation, while in urodeles (salamanders) and planarians (flatworms) persistent damage generally ends merely with functional repair. Injury response always entails attempts to repair the damage, but the fundamental differences probably lay in the coordination of such responses rather than in structural or species-specific capabilities” [17]. And evidence suggests that it is “individuation” or regenerative fields that direct and coordinate epimorphic regeneration in animals – a notion put forth by Conrad Hal Waddington in 1935 [18]. In 1938, Burr, Smith, and Strong found that bioelectric differences exist between cancer-susceptible and cancer-resistant mice, as well as that the onset of cancer is associated with a global deviation in an organism’s electrodynamic field [19]. Accordingly, we can state that cancer’s pathology is not merely the result of a random somatic mutation, and that it indeed consists of informational as well as molecular elements. Continuing, remodeling of the extracellular matrix is a major piece of limb regeneration and as a side note this process can be augmented with sufficient vitamin A [20]. Fibroblasts are prototypical cells that construct extracellular matrices and play central roles in both wound healing and tumor formation [21]. In salamanders, the dedifferentiation of fibroblasts (as well as some other cell types) after injury gives rise to what is known as a regeneration blastema (a mass of undifferentiated cells primed to repair lost tissue) that supplies the injured area with restorative cells [22]. In humans, the wound healing process can be divided into four phases: hemostasis, inflammation, proliferation, and tissue remodeling [23]. As the proliferation phase concludes in humans, certain fibroblasts (myofibroblasts) undergo apoptosis and are then cleared by macrophages [24]. In cancer, instead of being cleared away by macrophages, myofibroblasts remain in the diseased or injured region and become cancer-associated fibroblasts that are persistently activated by and promote the expansion of the developing tumor [25]. Now, earlier I mentioned the importance of reestablishing normalcy and health to the microenvironment of a tumor in the reversing of cancer because communication between neoplastic cells and their microenvironment drives tumor progression [26]. For corroboration, Bissell and Hines have explained that “the microenvironment can provide crucial signaling to maintain tissue architecture, inhibit cell growth and suppress or revert the malignant phenotype…incorrect signals from the microenvironment should lead to destabilization of tissue homeostasis and initiation and promotion of normal cells to malignancy” [27]. Salamanders masterfully create embryonic-like microenvironments at the site of regeneration after injury, and this not only ensures the correct reconstructing of harmed or missing tissue, but can also reprogram cancer cells (via signaling pathways) so that they revert to a normal phenotype [28]. Such reprogramming can also take place in the human body, and the homeobox genes (which code for proteins that help direct morphogenesis) that salamanders utilize in regeneration are also present in humans [29] [30]. Appropriate activation of beta-catenin and wnt signaling could trigger expression of the above genes that leads to commands and instructions for regeneration being given in Homo sapiens [31]. Alternatively, virus-mediated transduction could be used to extract functional homeobox genes from salamanders and insert them into human cells at sites requiring healing [32]. Immunomodulatory mesenchymal stem cells can be found throughout the body, and correct regeneration of human fingertips has been seen, in addition to the regenerative capacities displayed by human endometrium, kidney, liver, bladder, lung, gut, bone, skeletal muscle, and heart tissue [33] [34] [35]. So the commonly propagated idea that we uprights are largely incapable of true regeneration is incorrect. Digging deeper still, nerve dependence refers to the outgrowth of axons being necessary for the reconstituting of amputated limbs in animals [36]. In order for epimorphic regeneration to proceed rightly in some organisms, there must be a presence of regenerating nerves in the progenitor cell niche or microenvironment [37]. Interestingly, denervation of a neoplasm can stunt its expansion and suppress metastasis, suggesting nerve dependence in cancer [38]. Once more, there are parallels between the steps of regenerative healing and cancer, and it appears that the interaction of sprouting nerves with blastema and tumor cells is similar, meaning the innervation of proliferating tissue is capable of promoting either regeneration or tumorigenesis depending on the conditions of and signaling within the local microenvironment [39]. Evidently, neural facilitation of cancer progression includes the release of trophic factors by nerves toward tumor and stroma cells, the release of trophic factors by tumor cells beckoning nerve infiltration, activation of the stem cell compartment by neurotransmitters, and the supporting of neoangiogenesis [40]. So, in both regenerative blastemas and malignancies, there is bidirectional communication between sprouting nerves and proliferating cells [41]. Extrapolating a bit, we know that mitochondria can be transferred between cells in mammals, and that sabotaged mitochondrial respiration can be restored in neoplastic cells via the acquisition of mitochondria from other cells or the local microenvironment (which propagates tumor advancement) [42] [43]. Because of the known interaction between nerves, tumor cells, and microenvironments, and because nerve fibers reach into virtually every crevice of the body, the nervous system may not only aid in the apportioning of mitochondria to target tissues but also stand as an overseer in both regeneration and cancer. Coming to the last stop on our train ride, it is clear that mitochondrial dysfunction from free radicals or toxins, harmful electromagnetic radiation, hypoxia, viral infections, or abnormal mtDNA gene expression can induce a kind of metabolic reprogramming that mirrors the Warburg effect of cancer cells relying on glycolysis for their making of energy instead of oxidative phosphorylation and includes the acquisition of an invasive phenotype [44]. The arising of defects in mitochondria then may kickstart a signaling cascade that fosters tumor formation, whereas healthy mitochondria can suppress tumorigenesis [45]. As Seyfried et al. have expressed, “Any unspecific condition that damages a cell’s respiratory capacity but is not severe enough to kill the cell can potentially initiate the path to a malignant cancer” [46]. So the ability of cells to properly breathe is very important for a state of hypoxia or low tissue oxygen can initiate cell death via apoptosis (with the help of functional p53 proteins) or provoke adaptive responses that contribute to tumor formation [47]. And a hypoxic state encourages lactate production via glycolysis and subsequent tissue acidosis, which favors neoplasm progression and obstructs antitumor immunity (hypoxic and acidic extracellular spaces also propel invasion and metastasis) [48] [49]. Note that both chemotherapy and radiation therapy can damage mitochondrial respiration in healthy cells, negatively alter the tumor microenvironment, and encourage cancer recurrence [50] [51] [52] [53] [54] [55] [56]. Significant evidence supports the hypothesis that essentially all cancer hallmarks can be linked to impaired mitochondrial function and energy crafting [57]. In other words, cancer should be viewed primarily as a metabolic disease and not a genetic disease. Finally, the integrity of the nuclear genome leans heavily upon efficient and normal mitochondrial function as mitochondrial stress can engender DNA repair abnormalities and the upregulation of ATP making via glycolysis [58] [59] [60]. And this brings us full circle because the metabolic yields of high-rate glycolysis can destabilize the morphogenetic field of a microenvironment and contribute to inflammation and tumor advancement [61] [62]. We can equate ‘morphogenetic field’ to the term individuation field introduced earlier, and therefore conclude that individuation fields coordinate epimorphic regeneration when stable and healthy, yet advocate the manifestation of cancer when unstable and infirmed because of metabolic ill-health (resulting from such factors as hypoxia, acidosis, nutrient deficiencies, toxicity, and inflammation) with mitochondria and ATP production as the gateway. ATP is the energy of life, so when an organism becomes incapable of effectively and efficiently crafting ATP, the stage can be set for cancer’s appearance. Just as the salamander remains aligned with and ensconced in life by transmuting that which obstructs life, so too may we unhinder our ability to heal and forestall cancer by augmenting our transmutative faculty through eating alive foods, drinking alive water, shielding ourselves from discordant EMFs, detoxifying regularly, and staying positive in thought and feeling. The appropriate prevention and treatment of cancer then, lies not in the worsening of the body’s health and its power to make ATP, but in the improving of the body’s health and its power to make ATP – and this is why natural oncology experiences the success it does. References:
0 Comments
Your comment will be posted after it is approved.
Leave a Reply. |
AuthorDenton Coleman is an Exercise Physiologist and Medical Researcher. Archives
October 2023
Categories |

 RSS Feed
RSS Feed
